ZJU scientists identify the crucial role of a novel protein domain in photosystem II complex assembly and photoautotrophic growth in angiosperms
Recently, the research team led by MO Xiaorong at the Zhejiang University College of Life Sciences published an article titled “A novel protein domain is important for photosystem II complex assembly and photoautotrophic growth in angiosperms” in the journal Molecular Plant. They discovered a novel protein domain termed as the CLS (chlorotic lethal seedling) domain in TROL2, a second-type thylakoid rhodanese-like (TROL) protein in seed plant genomes. They identified the crucial role of this domain in photosystem II (PSII) complex assembly and photoautotrophic growth in angiosperms. Moreover, they proposed the origin of this domain during evolution.
TROL provides an anchor for leaf-type ferredoxin–NADP+ oxidoreductase. By comparing the functions of TROL1 and TROL2, MO Xiaorong et al. found that both of them were primarily expressed in rice leaves, localized to the chloroplast thylakoid membrane, and that they could interact with LFNR. However, TROL1 and TROL2 depletion in rice led to completely different phenotypes. The ostrol1 mutant resembled the wild type in phenotype, but the ostrol2 mutant displayed a chlorotic and lethal phenotype. Introducing a transgene of OsTROL2 driven by OsTROL2 promoter could restore the growth of the Ostrol2 mutant, but OsTROL1 driven by the same promoter failed to rescue the Ostrol2 mutant, suggesting that OsTROL2 is not just another LFNR anchor. Instead, its biochemical function is different from that of OsTROL1 and is vital for plant survival.
PSII is a multi-subunit protein complex vital to photosynthesis. MO Xiaorong et al. reported the function of TROL2 in PSII assembly, which differentiated OsTROL2 from OsTROL1. They also discovered the molecular mechanism of TROL2 in PSII assembly: TROL2 and LPA2 could be combined into compounds which could interact with PsbI, PsbL and PsbZ to promote PSII assembly.
Interestingly, they also identified a CLS domain in the TROL2 N terminus. This domain had only 45 amino acids, but it was both necessary and sufficient for TROL2 function in PSII assembly and photoautotrophic growth of angiosperms. Phylogenetic analyses revealed that TROL2 may have arisen from ancestral TROL1 via gene duplication before the emergence of seed plants and acquired the CLS domain via evolution of the sequence encoding the N-terminal part of the protein. In addition, the trol2 mutants in soybean, Arabidopsis and maize and soybean all exhibited seedling lethality, demonstrating that the TROL2 function was conserved in angiosperms. They also compared TROL in Marchantia polymorpha and TROL2 in angiosperms and proposed an intriguing question: Why is TROL2 essential for the photoautotrophic growth of angiosperms but unnecessary for the photoautotrophic growth of algae, bryophytes, or ferns?
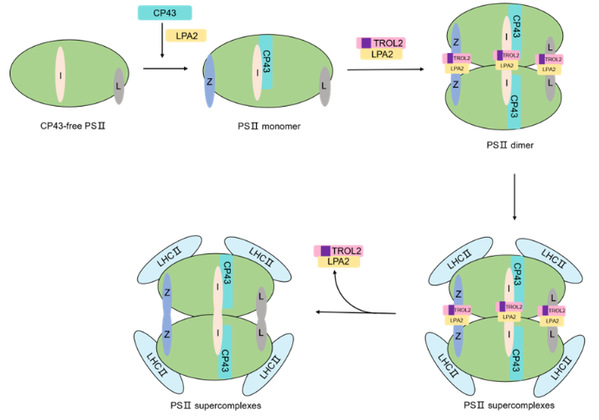
Model for the role of LPA2 and TROL2 in PSII complex assembly.
These findings provide a prime example of the generation of new genes via gene duplication and neofunctionalization during evolution. They are of immense theoretical significance for refining the molecular mechanism of photosynthetic complex assembly and the evolution of angiosperms.